
БНБ
"БСЭ" (95279)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Кора больших полушарий головного мозга
Определение "Кора больших полушарий головного мозга" в Большой Советской Энциклопедии
 |
| Кора головного мозга человека |
 |
| Пирамидная клетка коры головного мозга белой крысы |
 |
| Представительство чувствительных и двигательных функций тела |
 |
| Цитоархитектонические поля коры головного мозга человека |
Функциональные особенности коры обусловливаются упомянутым выше распределением нервных клеток и их связей по слоям и колонкам. На корковые нейроны возможна конвергенция (схождение) импульсов от различных органов чувств. Согласно современным представлениям, подобная конвергенция разнородных возбуждений - нейрофизиологический механизм интегративной деятельности головного мозга, т. е. анализа и синтеза ответной деятельности организма. Существенное значение имеет и то, что нейроны сведены в комплексы, по-видимому, реализующие результаты конвергенции возбуждений на отдельные нейроны. Одна из основных морфо-функциональных единиц коры - комплекс, называемый колонкой клеток, который проходит через все корковые слои и состоит из клеток, расположенных на одном перпендикуляре к поверхности коры. Клетки в колонке тесно связаны между собой и получают общую афферентную веточку из подкорки. Каждая колонка клеток отвечает за восприятие преимущественно одного вида чувствительности. Например, если в корковом конце кожного анализатора одна из колонок реагирует на прикосновение к коже, то другая - на движение конечности в суставе. В зрительном анализаторе функции восприятия зрительных образов также распределены по колонкам. Например, одна из колонок воспринимает движение предмета в горизонтальной плоскости, соседняя - в вертикальной и т. п.
Второй комплекс клеток новой коры - слой - ориентирован в горизонтальной плоскости. Полагают, что мелкоклеточные слои II и IV состоят в основном из воспринимающих элементов и являются «входами» в кору. Крупноклеточный слой V - выход из коры в подкорку, а среднеклеточный слой III - ассоциативный, связывающий между собой различные корковые зоны (см. рис. 1).
Локализация функций в коре характеризуется динамичностью в силу того, что, с одной стороны, имеются строго локализованные и пространственно отграниченные зоны коры, связанные с восприятием информации от определенного органа чувств, а с другой - кора является единым аппаратом, в котором отдельные структуры тесно связаны и в случае необходимости могут взаимозаменяться (т. н. пластичность корковых функций). Кроме того, в каждый данный момент корковые структуры (нейроны, поля, области) могут образовывать согласованно действующие комплексы, состав которых изменяется в зависимости от специфических и неспецифических стимулов, определяющих распределение торможения и возбуждения в коре. Наконец, существует тесная взаимозависимость между функциональным состоянием корковых зон и деятельностью подкорковых структур. Территории коры резко различаются по своим функциям. Большая часть древней коры входит в систему обонятельного анализатора. Старая и межуточная кора, будучи тесно связанными с древней корой как системами связей, так и эволюционно, не имеют прямого отношения к обонянию. Они входят в состав системы, ведающей регуляцией вегетативных реакций и эмоциональных состояний организма (см. Ретикулярная формация, Лимбическая система). Новая кора - совокупность конечных звеньев различных воспринимающих (сенсорных) систем (корковых концов анализаторов).
Принято выделять в зоне того или иного анализатора проекционные, или первичные, и вторичные, поля, а также третичные поля, или ассоциативные зоны. Первичные поля получают информацию, опосредованную через наименьшее количество переключений в подкорке (в зрительном бугре, или таламусе, промежуточного мозга). На этих полях как бы спроецирована поверхность периферических рецепторов (рис. 4). В свете современных данных, проекционные зоны нельзя рассматривать как устройства, воспринимающие раздражения «точку в точку». В этих зонах происходит восприятие определенных параметров объектов, т. е. создаются (интегрируются) образы, поскольку данные участки мозга отвечают на определенные изменения объектов, на их форму, ориентацию, скорость движения и т. п.
Кроме того, локализация функций в первичных зонах многократно дублируется по механизму, напоминающему голографию, когда каждый самый маленький участок запоминающего устройства содержит сведения о всём объекте. Поэтому достаточно сохранности небольшого участка первичного сенсорного поля, чтобы способность к восприятию почти полностью сохранилась. Вторичные поля получают проекции от органов чувств через дополнительные переключения в подкорке, что позволяет производить более сложный анализ того или иного образа. Наконец, третичные поля, или ассоциативные зоны, получают информацию от неспецифических подкорковых ядер, в которых суммируется информация от нескольких органов чувств, что позволяет анализировать и интегрировать тот или иной объект в ещё более абстрагированной и обобщённой форме. Эти области называются также зонами перекрытия анализаторов. Первичные и отчасти вторичные поля - возможный субстрат первой сигнальной системы, а третичные зоны (ассоциативные) - второй сигнальной системы, специфичной для человека (И. П. Павлов). Эти межанализаторные структуры определяют сложные формы мозговой деятельности, включающие и профессиональные навыки (нижнетеменная область), и мышление, планирование и целенаправленность действий (лобная область), и письменную и устную речь (нижняя лобная подобласть, височная, височно-теменно-затылочная и нижнетеменная области). Основные представители первичных зон в затылочной области - поле 17, где спроецирована сетчатка, в височной - поле 41, где спроецирован Кортиев орган, в прецентральной области - поле 4, где осуществляется проекция проприорецепторов в соответствии с расположением мускулатуры, в постцентральной - поля 3 и 1, где спроецированы экстерорецепторы в соответствии с их распределением в коже. Вторичные зоны представлены полями 8 и 6 (двигательный анализатор), 5 и 7 (кожный анализатор), 18 и 19 (зрительный анализатор), 22 (слуховой анализатор). Третичные зоны представлены обширными участками лобной области (поля 9, 10, 45, 44 и 46), нижнетеменной (поля 40 и 39), височно-теменно-затылочной (поле 37).
Корковые структуры играют первостепенную роль в обучении животных и человека. Однако образование некоторых простых условных рефлексов, главным образом с внутренних органов, может быть обеспечено подкорковыми механизмами. Эти рефлексы могут образовываться и на низших уровнях развития, когда ещё нет коры. Сложные условные рефлексы, лежащие в основе целостных актов поведения, требуют сохранности корковых структур и участия не только первичных зон корковых концов анализаторов, но и ассоциативных - третичных зон. Корковые структуры имеют прямое отношение и к механизмам памяти. Электрораздражение отдельных областей коры (например, височной) вызывает у людей сложные картины воспоминаний.
Характерная особенность деятельности коры - её спонтанная электрическая активность, регистрируемая в виде электроэнцефалограммы (ЭЭГ). В целом кора и её нейроны обладают ритмической активностью, которая отражает происходящие в них биохимические и биофизические процессы. Эта активность имеет разнообразную амплитуду и частоту (от 1 до 60 гц) и изменяется под влиянием различных факторов.
Ритмическая активность коры нерегулярна, однако можно по частоте потенциалов выделить несколько разных типов её (альфа-, бета-, дельта- и тета-ритмы). ЭЭГ претерпевает характерные изменения при многих физиологических и патологических состояниях (различных фазах сна, при опухолях, судорожных припадках и т. и.). Ритм, т. е. частота, и амплитуда биоэлектрических потенциалов коры задаются подкорковыми структурами, которые синхронизируют работу групп корковых нейронов, что и создаёт условия для их согласованных разрядов. Этот ритм связан с апикальными (верхушечными) дендритами пирамидных клеток. На ритмическую деятельность коры накладываются влияния, идущие от органов чувств. Так, вспышка света, щелчок или прикосновение к коже вызывают в соответствующих зонах т. н. первичный ответ, состоящий из ряда позитивных волн (отклонение электронного луча на экране осциллографа вниз) и негативной волны (отклонение луча вверх). Эти волны отражают деятельность структур данного участка коры и меняются в её различных слоях.
Филогенез и онтогенез коры. Кора - продукт длительного эволюционного развития, в процессе которого сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано (совершенствованием и взаимодействием различных воспринимающих и двигательных систем. У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты). В связи с неравномерностью роста отдельных структур новой коры её поверхность становится складчатой, покрываясь бороздами и извилинами. Совершенствование коры конечного мозга у млекопитающих неразрывно связано с эволюцией всех отделов центральной нервной системы. Этот процесс сопровождается интенсивным ростом прямых и обратных связей, соединяющих корковые и подкорковые структуры. Т. о., на более высоких этапах эволюции функции подкорковых образований начинают контролироваться корковыми структурами. Данное явление получило название кортиколизации функций. В результате кортиколизации ствол мозга образует с корковыми структурами единый комплекс, а повреждение коры на высших этапах эволюции приводит к нарушению жизненно важных функций организма. Наибольшие изменения и увеличение в процессе эволюции новой коры претерпевают ассоциативные зоны, в то время как первичные, сенсорные поля уменьшаются по относительной величине. Разрастание новой коры приводит к вытеснению старой и древней на нижнюю и срединную поверхности мозга.
Корковая пластинка появляется в процессе внутриутробного развития человека сравнительно рано - на 2-м месяце. Раньше всего выделяются нижние слои коры (VI-VII), затем - более высоко расположенные (V, IV, III и II; см. рис. 1). К 6 месяцам у эмбриона уже имеются все цитоархитектонические поля коры, свойственные взрослому человеку. После рождения в росте коры можно выделить три переломных этапа: на 2-3-м месяце жизни, в 2,5-3 года и в 7 лет. К последнему сроку цитоархитектоника коры полностью сформирована, хотя тела нейронов продолжают увеличиваться до 18 лет. Корковые зоны анализаторов завершают своё развитие раньше, и степень их увеличения меньше, чем у вторичных и третичных зон. Отмечается большое разнообразие в сроках созревания корковых структур у разных индивидуумов, что совпадает с разнообразием сроков созревания функциональных особенностей коры. Т. о., индивидуальное (онтогенез) и историческое (филогенез) развитие коры характеризуется сходными закономерностями.
Лит.: Орбели Л. А., Вопросы высшей нервной деятельности, М.- Л., 1949; Цитоархитектоника коры большого мозга человека. Сб. ст., М., 1949; Филимонов И. Н., Сравнительная анатомия коры большого мозга млекопитающих, М., 1949; Павлов И. П., Двадцатилетний опыт объективного изучения высшей нервной деятельности животных, Полн. собр. соч., 2 изд., т. 3, кн. 1-2, М., 1951; Брейзье М., Электрическая активность нервной системы, пер. с англ., М., 1955; Сепп Е. К., История развития нервной системы позвоночных, 2 изд., М., 1959; Лурия А. Р., Высшие корковые функции человека и их нарушения при локальных поражениях мозга, М., 1962; Воронин Л. Г., Курс лекций по физиологии высшей нервной деятельности, М., 1965; Поляков Г. И., О принципах нейронной организации мозга, М., 1965; Корковая регуляция деятельности подкорковых образований головного мозга. Сб. ст., Тб., 1968; Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968; Беритов И. С., Структура и функции коры большого мозга, М., 1969.
Л. Г. Воронин.
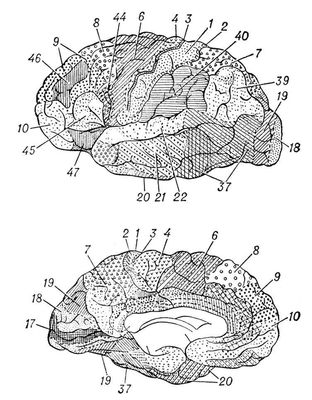
Рис. 3. Карта цитоархитектонических полей коры головного мозга человека: А - наружная поверхность полушария, Б - внутренняя поверхность полушария. Номерами и различной штриховкой обозначены цитоархитектонические поля коры.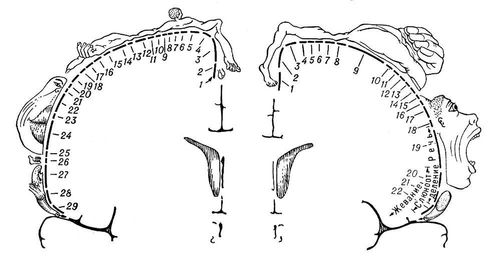
Рис. 4. Представительство чувствительных функций тела в задней центральной извилине (А) и двигательных функций - в передней центральной извилине (Б). А: 1 - половые органы; 2 - пальцы; 3 - ступня; 4 - голень; 5 - бедро; 6 - туловище; 7 - шея; 8 - голова; 9 - плечо; 10 - рука; 11 - локоть; 12 - предплечье; 13 - запястье; 14 - кисть; 15 - мизинец; 16 - безымянный палец; 17 - средний палец; 18 - указательный палец; 19 - большой палец; 20 - глаз; 21 - нос; 22 - лицо; 23 - верхняя губа; 24 - губы; 25 - нижняя губа; 26 - зубы, дёсны и челюсть; 27 - язык; 28 - глотка; 29 - внутренние органы. Б: 1 - пальцы; 2 - лодыжка; 3 - колено; 4 - бедро; 5 - туловище; 6 - плечо; 7 - локоть; 8 - запястье; 9 - кисть; 10 - мизинец; 11 - безымянный палец; 12 - средний палец; 13 - указательный палец; 14 - большой палец; 15 - шея; 16 - бровь; 17 - веко и глазное яблоко; 18 - лицо; 19 - губы; 20 - челюсть; 21 - язык; 22 - глотание. Размеры частей тела, изображенного на рисунке, соответствуют представительству двигательных и чувствительных функций организма в передней и задней центральных извилинах коры.
I - зональный слой, II - наружный зернистый слой, III - пирамидный слой, IV - внутренний зернистый слой, V - ганглионарный слой, VI - слой треугольных клеток, VII - слой веретеновидных клеток. А - нейронное строение, Б - цитоархитектоника, В - волоконная структура." href="/a_pictures/18/10/221347173.jpg">I - зональный слой, II - наружный зернистый слой, III - пирамидный слой, IV - внутренний зернистый слой, V - ганглионарный слой, VI - слой треугольных клеток, VII - слой веретеновидных клеток. А - нейронное строение, Б - цитоархитектоника, В - волоконная структура." src="a_pictures/18/10/th_221347173.jpg">
Рис. 1. Схема строения коры головного мозга человека: I - зональный слой, II - наружный зернистый слой, III - пирамидный слой, IV - внутренний зернистый слой, V - ганглионарный слой, VI - слой треугольных клеток, VII - слой веретеновидных клеток. А - нейронное строение, Б - цитоархитектоника, В - волоконная структура.
| "БСЭ" >> "К" >> "КО" >> "КОР" >> "КОРА" |
Статья про "Кора больших полушарий головного мозга" в Большой Советской Энциклопедии была прочитана 1078 раз
| Коптим скумбрию в коробке |
| Луковый соус |
TOP 20
- Лемке Михаил Константинович
- Сульфгидрильные группы
- «Казарменный коммунизм»
- Японское море
- Периодическая система элементов
- Объединённая партия гаитянских коммунистов
- Глициния
- Башкирская Автономная Советская Социалистическая Республика
- Иммунитет (историч.)
- Андаманское море
- Сенсуализм
- Балкано-кавказская раса
- «Сообразительный»
- Навигация (морск.)
- Звёздная астрономия
- Ямполь (пос. гор. типа в Донецкой обл.)
- Кульчицкая Елена Львовна
- Мандельштама - Бриллюэна рассеяние
- Театральные учебные заведения
- Электрическая постоянная