
БНБ
"БСЭ" (95279)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Хромосомы
Определение "Хромосомы" в Большой Советской Энциклопедии
 |
| Мейоз (схема) |
 |
| Митоз и изменение внешнего вида хромосом (схемы) |
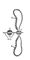 |
| Структура хромосомы и хромомеры |
 |
| Хромосома в митозе и мейозе |
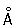 , называемые РНП-гранулами, интергранулами и перихроматиновыми гранулами. Эти частицы представляют собой форму упаковки РНК, синтезированной на Хромосомы и соединённой с белком, и служат для завершения образования информационной РНК и переноса её в цитоплазму.
, называемые РНП-гранулами, интергранулами и перихроматиновыми гранулами. Эти частицы представляют собой форму упаковки РНК, синтезированной на Хромосомы и соединённой с белком, и служат для завершения образования информационной РНК и переноса её в цитоплазму.
Для изучения интерфазных Хромосомы используют либо биохимические методы выделения вещества Хромосомы — хроматина и разделения его на эухроматин и гетерохроматин, либо электронно-микроскопическое исследование интактных ядер и изолированного хроматина; как модели интерфазных Хромосомы используют гигантские Хромосомы типа ламповых щёток из ооцитов животных и многонитчатые (политенные) Хромосомы двукрылых. В Хромосомы типа ламповых щёток неактивные участки имеют вид плотно упакованных структур — хромомер (рис. 2, 3), которые обнаруживаются и в Хромосомы соматических клеток, особенно в профазе митоза, и рассматриваются как характерные морфологические, а возможно и функциональные, единицы Хромосомы В участках Хромосомы, активно синтезирующих РНК, хромомеры раскручиваются и образуют боковые петли, в которых молекулы РНК, соединяясь с белком, образуют рибонуклеопротеиды (РНП) — частицы, представляющие собой форму упаковки генных продуктов и различающиеся в отдельных боковых петлях по размерам и морфологическим признакам. В политенных Хромосомы, возникающих в тканях двукрылых и некоторых растений за счёт многократной репликации (удвоения) исходной Хромосомы без последующего расхождения дочерних Хромосомы, неактивные участки имеют форму дисков, а активные образуют вздутия — пуфы. В пуфах, так же как и в Хромосомы типа ламповых щёток, содержатся частицы РНП диаметром 200—500 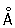 . Электронно-микроскопические и биохимические исследования показали, что и в хроматине, выделенном из клеток, и в интактных ядрах, и в гигантских Хромосомы основной структурной единицей является дезоксирибонуклеопротеидная нить (ДНП) диаметром,100—200
. Электронно-микроскопические и биохимические исследования показали, что и в хроматине, выделенном из клеток, и в интактных ядрах, и в гигантских Хромосомы основной структурной единицей является дезоксирибонуклеопротеидная нить (ДНП) диаметром,100—200 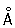 .
.
Изучение политенных Хромосомы в разных тканях и на разных стадиях развития двукрылых показало, что число и набор активных пуфов имеют тканевую и видовую специфичность. Это значит, что хотя все клетки многоклеточного организма имеют одинаковый набор генов, линейно расположенных в каждой Хромосомы, набор активных и неактивных в синтезе РНК участков Хромосомы различается в каждом типе клеток и на разных стадиях развития, т. е. один и тот же участок находится в одних тканях в эухроматическом, в других — в гетерохроматическом состоянии. Отдельные участки Хромосомы находятся в гетерохроматическом состоянии в интерфазе разных типов клеток; как правило, они отличаются присутствием высокоповторяющихся последовательностей ДНК. Постоянно функционирующим в интерфазе всех типов клеток является ядрышковый организатор — участок Хромосомы, где сосредоточены гены рибосомной РНК. В этой области формируется ядрышко, которое долго считали самостоятельным органоидом клетки. Оно является местом формирования предшественников рибосом.
Хромосомы в интерфазном ядре отделены от цитоплазмы ядерной мембраной; многими участками (прежде всего, теломерами и центромерами) они соединены с ней, благодаря чему, как полагают, каждая Хромосомы занимает в ядре определённое место. При подготовке клетки к делению в интерфазе происходит удвоение Хромосомы Каждая Хромосомы строит свою копию на основе полуконсервативной репликации ДНК. Особенностью Хромосомы эукариот является существование многих точек начала и завершения репликации (у прокариот лишь одна точка начала и одна точка завершения репликации). Этим обеспечивается возможность неодновременной репликации разных участков Хромосомы в ходе синтетического периода и регуляция активности Хромосомы
Хромосомы в период митоза и мейоза. При переходе клетки к делению синтез ДНК и РНК в Хромосомы прекращается, Хромосомы приобретают всё более плотную упаковку (например, в одной Хромосомы человека цепочка ДНК длиной 160 мм укладывается в объёме всего 0,5´10 мкм), ядерная мембрана разрушается и Хромосомы выстраиваются на экваторе клетки. В этот период они наиболее доступны для наблюдения и изучения их морфологии. Основная структурная единица метафазных Хромосомы, так же как и интерфазных, — нить ДНП диаметром 100—200 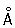 , уложенная в плотную спираль. Некоторые авторы обнаруживают, что нити диаметром 100—200
, уложенная в плотную спираль. Некоторые авторы обнаруживают, что нити диаметром 100—200 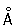 образуют структуры второго уровня укладки — нити диаметром около 2000
образуют структуры второго уровня укладки — нити диаметром около 2000 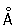 , которые и формируют тело метафазной Хромосомы Каждая метафазная Хромосомы состоит из хроматид (рис. 3, 1), образовавшихся в результате репликации исходной интерфазной Хромосомы Использование меченых и модифицированных предшественников ДНК позволило четко различать в Хромосомы, находящейся в метафазе митоза, дифференциально окрашенные хроматиды, благодаря чему было установлено, что при репликации Хромосомы нередко происходит обмен участками между сестринскими хроматидами (кроссинговер). В классической цитологии придавалось большое значение матриксу метафазной Хромосомы, его считали обязательным компонентом, в который погружены спирализованные хромонемы. Современные цитологи рассматривают матрикс метафазных Хромосомы как остаточный материал разрушающегося ядрышка; часто он вовсе не обнаруживается.
, которые и формируют тело метафазной Хромосомы Каждая метафазная Хромосомы состоит из хроматид (рис. 3, 1), образовавшихся в результате репликации исходной интерфазной Хромосомы Использование меченых и модифицированных предшественников ДНК позволило четко различать в Хромосомы, находящейся в метафазе митоза, дифференциально окрашенные хроматиды, благодаря чему было установлено, что при репликации Хромосомы нередко происходит обмен участками между сестринскими хроматидами (кроссинговер). В классической цитологии придавалось большое значение матриксу метафазной Хромосомы, его считали обязательным компонентом, в который погружены спирализованные хромонемы. Современные цитологи рассматривают матрикс метафазных Хромосомы как остаточный материал разрушающегося ядрышка; часто он вовсе не обнаруживается.
Формирование половых клеток у животных и растений сопровождается особым типом их деления — мейозом, и мейотические Хромосомы имеют ряд особенностей по сравнению с митотическими. Прежде всего, при мейозе дочерние клетки получают вдвое уменьшенное число Хромосомы (при митозе оно сохраняется одинаковым), что достигается благодаря конъюгации гомологичных Хромосомы в профазе мейоза и двумя последовательными делениями клетки при одной репликации ДНК (подробнее см. Мейоз). Кроме того, у мейотические Хромосомы отмечаются временный перерыв профазы мейоза и возвращение их к интерфазному состоянию, когда Хромосомы начинают активно синтезировать РНК. В этом периоде у большинства изученных животных организмов наблюдаются Хромосомы типа ламповых щёток (рис. 4). Наконец, Хромосомы в метафазе мейоза отличаются более плотной упаковкой.
Несмотря на огромное число исследований, посвященных Хромосомы, изучение их структурной и функциональной организации продолжает оставаться одним из самых актуальных направлений современной биологии. Хромосомы выполняют в клетке сложнейшие функции и имеют весьма сложную организацию, трудно поддающуюся изучению. Огромные успехи в понимании молекулярных основ строения Хромосомы достигнуты в 60—70-е гг. 20 в. благодаря развитию молекулярной генетики. Эти успехи блестяще подтвердили основные положения хромосомной теории наследственности, углубив и развив их.
Лит.: Вильсон Э., Клетка и ее роль в развитии и наследственности, пер. с англ., т. 1—2, М. — Л., 1936—40; Кольцов Н. К., Организация клетки, М. — Л., 1936; Прокофьева-Бельговская А. А., Строение хромосомы, в кн.: Ионизирующие излучения и наследственность, М., 1960 (Итоги науки. Биологические науки, в. 3); Кикнадзе И. И., Функциональная организация хромосом, Л., 1972; Де Робертис Э., Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1973; Левитский Г. А., Цитология растений. Избр. труды, М., 1976; Darlington С. D., Recent advances in cytology, 2 ed., L., 1937; Geitler L., Chromosomenbau, B., 1938 (Protoplasma-Monographien, Bd 14); Ris Н., Kubai D. F., Chromosome structure, «Annual Review of Genetics», 1970, v. 4, p. 236—94; Handbook of molecular cytology, ed. by Lima-de-Faria A., Amst. — L., 1969; Chromosome structure and function, N. Y., 1974.
И. И. Кикнадзе.
| "БСЭ" >> "Х" >> "ХР" >> "ХРО" |
Статья про "Хромосомы" в Большой Советской Энциклопедии была прочитана 1471 раз
| Бургер двойного помола |
| Салат тофу |
TOP 20
- Лемке Михаил Константинович
- Сульфгидрильные группы
- «Казарменный коммунизм»
- Японское море
- Периодическая система элементов
- Глициния
- Башкирская Автономная Советская Социалистическая Республика
- Иммунитет (историч.)
- Андаманское море
- Сенсуализм
- «Сообразительный»
- Балкано-кавказская раса
- Навигация (морск.)
- Звёздная астрономия
- Ямполь (пос. гор. типа в Донецкой обл.)
- Кульчицкая Елена Львовна
- Мандельштама - Бриллюэна рассеяние
- Театральные учебные заведения
- Электрическая постоянная
- Энсор Джеймс