
БНБ
"БСЭ" (95279)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Хромосомная теория наследственности
Определение "Хромосомная теория наследственности" в Большой Советской Энциклопедии
 |
| Хромосомная теория наследственности. Рис. |
В 1902 У. Сеттон в США, обративший внимание на параллелизм в поведении хромосом и менделевских т. н. «наследственных факторов», и Т. Бовери в Германии выдвинули хромосомную гипотезу наследственности, согласно которой менделевские наследственные факторы (название впоследствии генами) локализованы в хромосомах. Первые подтверждения этой гипотезы были получены при изучении генетического механизма определения пола у животных, когда было выяснено, что в основе этого механизма лежит распределение половых хромосом среди потомков. Дальнейшее обоснование Хромосомная теория наследственности принадлежит американскому генетику Т. Х. Моргану, который заметил, что передача некоторых генов (например, гена, обусловливающего белоглазие у самок дрозофилы при скрещивании с красноглазыми самцами) связана с передачей половой Х-хромосомы, т. е. что наследуются признаки, сцепленные с полом (у человека известно несколько десятков таких признаков, в том числе некоторые наследственные дефекты - дальтонизм, гемофилия и др.).
Доказательство Хромосомная теория наследственности было получено в 1913 американским генетиком К. Бриджесом, открывшим нерасхождение хромосом в процессе мейоза у самок дрозофилы и отметившим, что нарушение в распределении половых хромосом сопровождается изменениями в наследовании признаков, сцепленных с полом.
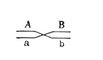
С развитием Хромосомная теория наследственности было установлено, что гены, расположенные в одной хромосоме, составляют одну группу сцепления (см. Сцепление генов) и должны наследоваться совместно; число групп сцепления равно числу пар хромосом, постоянному для каждого вида организмов (см. Кариотип); признаки, зависящие от сцепленных генов, также наследуются совместно. Вследствие этого закон независимого комбинирования признаков (см. Менделя законы) должен иметь ограниченное применение; независимо должны наследоваться признаки, гены которых расположены в разных (негомологичных) хромосомах. Явление неполного сцепления генов (когда наряду с родительскими сочетаниями признаков в потомстве от скрещиваний обнаруживаются и новые, рекомбинантные, их сочетания) было подробно исследовано Морганом и его сотрудниками (А. Г. Стёртевантом и др.) и послужило обоснованием линейного расположения генов в хромосомах. Морган предположил, что сцепленные гены гомологичных хромосом, находящиеся у родителей в сочетаниях 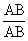 и
и 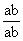 , в мейозе у гетерозиготной формы ®
, в мейозе у гетерозиготной формы ® 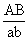 могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами
могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами 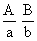 и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или кроссинговером, была доказана в 1933 нем, учёным К. Штерномв опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток - с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения генетических карт хромосом. В 30-х гг. 20 в. Ф. Добржанский показал, что порядок размещения генов на генетических и цитологических картах хромосом совпадает.
и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или кроссинговером, была доказана в 1933 нем, учёным К. Штерномв опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток - с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения генетических карт хромосом. В 30-х гг. 20 в. Ф. Добржанский показал, что порядок размещения генов на генетических и цитологических картах хромосом совпадает.
Согласно представлениям школы Моргана, гены являются дискретными и далее неделимыми носителями наследственной информации. Однако открытие в 1925 советскими учёными Г. А. Надсоном и Г. С. Филипповым, а в 1927 американским учёным Г. Мёллером влияния рентгеновских лучей на возникновение наследственных изменений (мутаций) у дрозофилы, а также применение рентгеновских лучей для ускорения мутационного процесса у дрозофилы позволили советским учёным А. С. Серебровскому, Н. П. Дубинину и др. сформулировать в 1928-30 представления о делимости гена на более мелкие единицы, расположенные в линейной последовательности и способные к мутационным изменениям. В 1957 эти представления были доказаны работой американского учёного С. Бензера с бактериофагом Т4. Использование рентгеновских лучей для стимулирования хромосомных перестроек позволило Н. П. Дубинину и Б. Н. Сидорову обнаружить в 1934 эффект положения гена (открытый в 1925 Стёртевантом), т. е. зависимость проявления гена от места расположения его на хромосоме. Возникло представление о единстве дискретности и непрерывности в строении хромосомы.
Хромосомная теория наследственности развивается в направлении углубления знаний об универсальных носителях наследственной информации - молекулах дезоксирибонуклеиновой кислоты (ДНК). Установлено, что непрерывная последовательность пуриновых и пиримидиновых оснований вдоль цепи ДНК образует гены, межгенные интервалы, знаки начала и конца считывания информации в пределах гена; определяет наследственный характер синтеза специфических белков клетки и, следовательно, наследственный характер обмена веществ. ДНК составляет материальную основу группы сцепления у бактерий и многих вирусов (у некоторых вирусов носителем наследственной информации является рибонуклеиновая кислота); молекулы ДНК, входящие в состав митохондрий, пластид и др. органоидов клетки, служат материальными носителями цитоплазматической наследственности.
Хромосомная теория наследственности, объясняя закономерности наследования признаков у животных и растительных организмов, играет важную роль в с.-х. науке и практике. Она вооружает селекционеров методами выведения пород животных и сортов растений с заданными свойствами. Некоторые положения Хромосомная теория наследственности позволяют более рационально вести с.-х. производство. Так, явление сцепленного с полом наследования ряда признаков у с.-х. животных позволило до изобретения методов искусственного регулирования пола у тутового шелкопряда выбраковывать коконы менее продуктивного пола, до разработки способа разделения цыплят по полу исследованием клоаки - отбраковывать петушков и т.п. Важнейшее значение для повышения урожайности многих с.-х. культур имеет использование полиплоидии. На знании закономерностей хромосомных перестроек основывается изучение наследственных заболеваний человека.
Лит.: Морган Т. Г., Структурные основы наследственности, пер. с англ., М.-П., 1924; его же, Избранные работы по генетике, пер, с англ., М.-Л., 1937; Актуальные вопросы современной генетики, М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Классики советской генетики. [Сб. ст.], Л., 1968.
С. Г. Инге-Вечтомов.
| "БСЭ" >> "Х" >> "ХР" >> "ХРО" |
Статья про "Хромосомная теория наследственности" в Большой Советской Энциклопедии была прочитана 1383 раз
| Пицца в сковороде |
| Луковый соус |
TOP 20
- Лемке Михаил Константинович
- Сульфгидрильные группы
- «Казарменный коммунизм»
- Японское море
- Периодическая система элементов
- Башкирская Автономная Советская Социалистическая Республика
- Объединённая партия гаитянских коммунистов
- Глициния
- Иммунитет (историч.)
- Андаманское море
- Сенсуализм
- Балкано-кавказская раса
- «Сообразительный»
- Звёздная астрономия
- Навигация (морск.)
- Ямполь (пос. гор. типа в Донецкой обл.)
- Мандельштама - Бриллюэна рассеяние
- Театральные учебные заведения
- Кульчицкая Елена Львовна
- Электрическая постоянная