
БНБ
"БСЭ" (95279)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Окисление биологическое
Определение "Окисление биологическое" в Большой Советской Энциклопедии
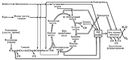 |
| Образование АТФ при хемоорганотрофном типе энергетического обмена |
Окисление биологическое в клетках связано с передачей т. н. восстанавливающих эквивалентов (ВЭ) — атомов водорода или электронов — от одного соединения — донора, к другому — акцептору. У аэробов — большинства животных, растений и многих микроорганизмов — конечным акцептором ВЭ служит кислород. Поставщиками ВЭ могут быть как органические, так и неорганические вещества (см. таблицу).
Классификация организмов по источнику энергии и восстанавливающих эквивалентов
| Источник энергии< | Окисляемое соединение (поставщик восстанавливающих эквивалентов) | Примеры |
| Фотолитотрофы Фотоорганотрофы Хемолитотрофы
Хемоорганотрофы | Свет Свет Реакции окисления
Реакции окисления | Неорганические соединения (Н2О, H2S, S) Органические соединения Неорганические соединения (H2, S, H2S, NH3, Fe2+)
Органические соединения | Зелёные клетки высших растений, синезелёные водоросли, фотосинтезирующие бактерии Несерные пурпурные бактерии Водородные, серные, денитрифицирующие бактерии, железобактерии
Животные, большинство микроорганизмов, нефотосинтезирующие клетки растений |
Основной путь использования энергии, освобождающейся при Окисление биологическое, — накопление её в молекулах аденозинтрифосфорной кислоты (АТФ) и др. макроэргических соединений. Окисление биологическое, сопровождающееся синтезом АТФ из аденозиндифосфорной кислоты (АДФ) и неорганического фосфата, происходит при гликолизе, окислении a-кетоглутаровой кислоты и при переносе ВЭ в цепи окислительных (дыхательных) ферментов, обычно называют окислительным фосфорилированием (см. схему).
В процессе дыхания углеводы, жиры и белки подвергаются многоступенчатому окислению, которое приводит к восстановлению основных поставщиков ВЭ для дыхательных флавинов, никотинамидадениндинуклеотида (НАД), никотинамидадениндинуклеотидфосфата (НАДФ) и липоевой кислоты. Восстановление этих соединений в значительной мере осуществляется в трикарбоновых кислот цикле, которым завершаются основные пути окислительного расщепления углеводов (оно начинается с гликолиза), жиров и аминокислот. Помимо цикла трикарбоновых кислот, некоторое количество восстановленных коферментов — ФАД (флавинадениндинуклеотида) и НАД — образуется при окислении жирных кислот, а также при окислительном дезаминировании глутаминовой кислоты (НАД) и в пентозофосфатном цикле (восстановленный НАДФ).
Соотношение и локализация различных механизмов Окисление биологическое В расчёте на 1 молекулу глюкозы гликолиз даёт 2 молекулы АТФ, а фосфорилирование в дыхательной цепи — 34 молекулы АТФ. Гликолиз, цикл трикарбоновых кислот и дыхательная цепь функционируют, по-видимому, в клетках всех эукариотов. Окисление жирных кислот у позвоночных поставляет половину энергии, потребляемой печенью, почками, мышцей сердца и покоящимися скелетными мышцами; в клетках мозга оно практически не происходит. Окисление по пентозофосфатному пути активно в печени и лактирующих молочных железах, но незначительно в сердечной и скелетных мышцах.
В жидкой фазе цитоплазмы растворены все ферменты гликолиза. Внутренние мембраны митохондрий, мембраны хлоропластов (тилакоидов) и клеточные мембраны бактерий содержат фосфорилирующие цепи переноса электронов. В матриксе митохондрий локализовано окисление жирных кислот, ферменты цикла трикарбоновых кислот и глутаматдегидрогеназа. Во внутренней мембране митохондрий находятся ферменты, окисляющие янтарную и b-оксимасляную кислоты, во внешней — ферменты, участвующие в обмене аминокислот: моноаминоксидаза и кинуренингидроксилаза. В особых органоидах клетки, т. н. пероксисомах, или микротельцах, вклад которых в суммарное поглощение О2 может достигать в печени 20%, находится флавиновая оксидаза, окисляющая аминокислоты, гликолевую кислоту и др. субстраты с образованием перекиси водорода, которая затем разлагается каталазой или используется пероксидазами в реакциях окисления. В мембранах эндоплазматической сети клетки локализованы гидроксилазы и оксигеназы, организованные в короткие нефосфорилирующие цепи переноса электронов.
Окислительные реакции не всегда сопровождаются накоплением энергии; в ряде случаев они несут функции превращения веществ (например, окисление при образовании жёлчных кислот, стероидных гормонов, на путях превращения аминокислот и др.). При окислении происходит обезвреживание чужеродных и ядовитых для организма веществ (ароматических соединений, недоокисленных продуктов дыхания и др.). Окисление биологическое, не сопряжённое с накоплением энергии, называется свободным окислением. Его энергетический эффект — образование тепла. По-видимому, система переноса электронов, осуществляющая окислительное фосфорилирование, способна переключаться на свободное окисление при увеличении потребности организма в тепле (у гомойотермных животных).
Механизм использования энергии окисления. Долгое время оставался неясным вопрос о механизме преобразования энергии, освобождающейся при переносе ВЭ по цепи окислительных ферментов. Согласно т. н. хемиосмотической теории, развитой в 60-х гг. 20 в. (английский биохимик П. Митчелл и др.), энергия сначала используется для создания электрического поля («+» с одной стороны мембраны и «–» с другой) и разности концентраций ионов Н+ по разные стороны мембраны. Оба фактора (электрическое поле и разность концентраций) могут служить движущей силой для действия фермента АТФ-синтетазы, осуществляющей синтез АТФ. Часть энергии поля может быть прямо использована клеткой для переноса ионов через мембрану, восстановление переносчиков электронов, образования тепла без промежуточного участия АТФ.
Эволюция энергообеспечения в живой природе. Древнейшие организмы, как полагают, существовали в первичной бескислородной атмосфере Земли и были анаэробами и гетеротрофными организмами. Обеспечение клеток энергией шло за счёт процессов типа гликолиза. Возможно, существовал механизм окисления, известный у некоторых современных микроорганизмов: ВЭ передаются через дыхательную цепь на нитрат (NO–3) или на сульфат (SO– –4). Принципиально важным этапом оказалось возникновение у древних одноклеточных организмов механизма фотосинтеза, с которым связывают появление кислорода в атмосфере Земли. В результате стало возможным использование O2, обладающего высоким окислительно-восстановительным потенциалом, в качестве конечного акцептора электронов в дыхательной цепи. Реализация этой возможности произошла при появлении специального фермента — цитохромоксидазы, восстанавливающей О2, и привела к возникновению биохимического дыхательного аппарата современного типа. Обеспечение энергией у всех аэробов (их клетки содержат митохондрии) основано на таком дыхании. Вместе с тем клетки сохранили ферментный аппарат гликолиза. Образуемая в ходе последнего пировиноградная кислота окисляется далее в цикле трикарбоновых кислот, который, в свою очередь, питает дыхательную цепь электронами. Т. о., эволюция энергетического обмена шла, по-видимому, по пути использования и надстройки уже имевшихся ранее механизмов энергообеспечения. Наличие в клетках ныне существующих организмов биохимических систем гликолиза (в цитоплазме), дыхания (в митохондриях), фотосинтеза (в хлоропластах), а также поразительное сходство механизмов превращения энергии в этих органеллах и в микроорганизмах нередко рассматривают как свидетельство возможного происхождения хлоропластов и митохондрий от древних микроорганизмов-симбионтов. См. также Аденозинфосфорные кислоты, Биоэнергетика, Брожение, Дыхание, Митохондрии, Фотосинтез и лит. при этих статьях.
Лит.: Ленингер А., Превращение энергии в клетке, в кн.: Живая клетка, пер. с англ., М., 1962; Скулачев В. П., Аккумуляция энергии в клетке, М., 1969; его же, Трансформация энергии в биомембранах, М., 1972; Малер Г. и Кордес Ю., Основы биологической химии, пер. с англ., М., 1970, гл. 15; Леви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971, гл. 12; Ясайтис А. А., Превращение энергии в митохондриях, М., 1973; Ленинджер А., Биохимия, пер. с англ., М., 1974.
С. А. Остроумов.
| "БСЭ" >> "О" >> "ОК" >> "ОКИ" |
Статья про "Окисление биологическое" в Большой Советской Энциклопедии была прочитана 1159 раз
| Коптим скумбрию в коробке |
| Коптим скумбрию дома в коробке |
TOP 20
- Лемке Михаил Константинович
- Сульфгидрильные группы
- «Казарменный коммунизм»
- Японское море
- Периодическая система элементов
- Объединённая партия гаитянских коммунистов
- Глициния
- Башкирская Автономная Советская Социалистическая Республика
- Иммунитет (историч.)
- Андаманское море
- Сенсуализм
- Балкано-кавказская раса
- «Сообразительный»
- Навигация (морск.)
- Звёздная астрономия
- Ямполь (пос. гор. типа в Донецкой обл.)
- Мандельштама - Бриллюэна рассеяние
- Кульчицкая Елена Львовна
- Театральные учебные заведения
- Электрическая постоянная